真菌病原体已经进化出专门的侵染相关结构,以穿透植物坚硬的外层,引起病害。这些侵染结构包括附着胞等,是一些最严重的作物病害的重要决定因素,包括谷物锈病和白粉病,但它们的生物学特性尚不清楚。形成这些结构需要重大的形态发生变化和信号转导事件的重塑,包括磷酸化。其中一些事件由主要的调节因子控制,如丝裂原活化蛋白激酶(MAPKs)。水稻稻瘟病菌Magnaporthe oryzae,每年破坏足够养活6000万人的稻米,发育出一个黑色素色素沉着的附着胞,产生高达8.0 MPa的巨大渗透压,使病原体能够破裂坚硬的水稻叶片表皮。M. oryzae中附着胞形态发生需要一个MAPK信号通路,其中病原性丝裂原活化蛋白激酶1(Pmk1)MAPK是核心组成部分。Pmk1的重要性通过以下事实得到证明:Δpmk1突变体无法形成附着胞,无法引起稻瘟病,而使用模拟敏感突变体的条件性激酶失活则可以防止真菌在植物组织中的侵袭性生长。全转录分析揭示了Pmk1在附着胞发育期间调控49%的M. oryzae转录组,突出了其重要性和侵染相关形态发生的复杂性。Pmk1级联的组成部分主要是基于它们在已知的酿酒酵母Fus3/Kss1途径中的对应蛋白来鉴定的,该途径需要用于信息素信号和侵袭性生长。例如,上游激酶Mst11和Mst7分别基于它们与酿酒酵母MAPKKK Ste11和MAPKK Ste7的同源性进行了功能表征。同样,通过与Ste50的同源性鉴定了适配蛋白Mst50,并显示它控制着附着胞形成期间Mst11-Mst7-Pmk1 MAPK模块的活性。Pmk1下游的少数相互作用因子也已被鉴定,包括转录因子Mst12、Hox7和Slf1,以及与Pmk1相互作用的克隆Pic5。然而,这些下游Pmk1信号组分如何调节稻瘟病感染的分子机制仍然未知。

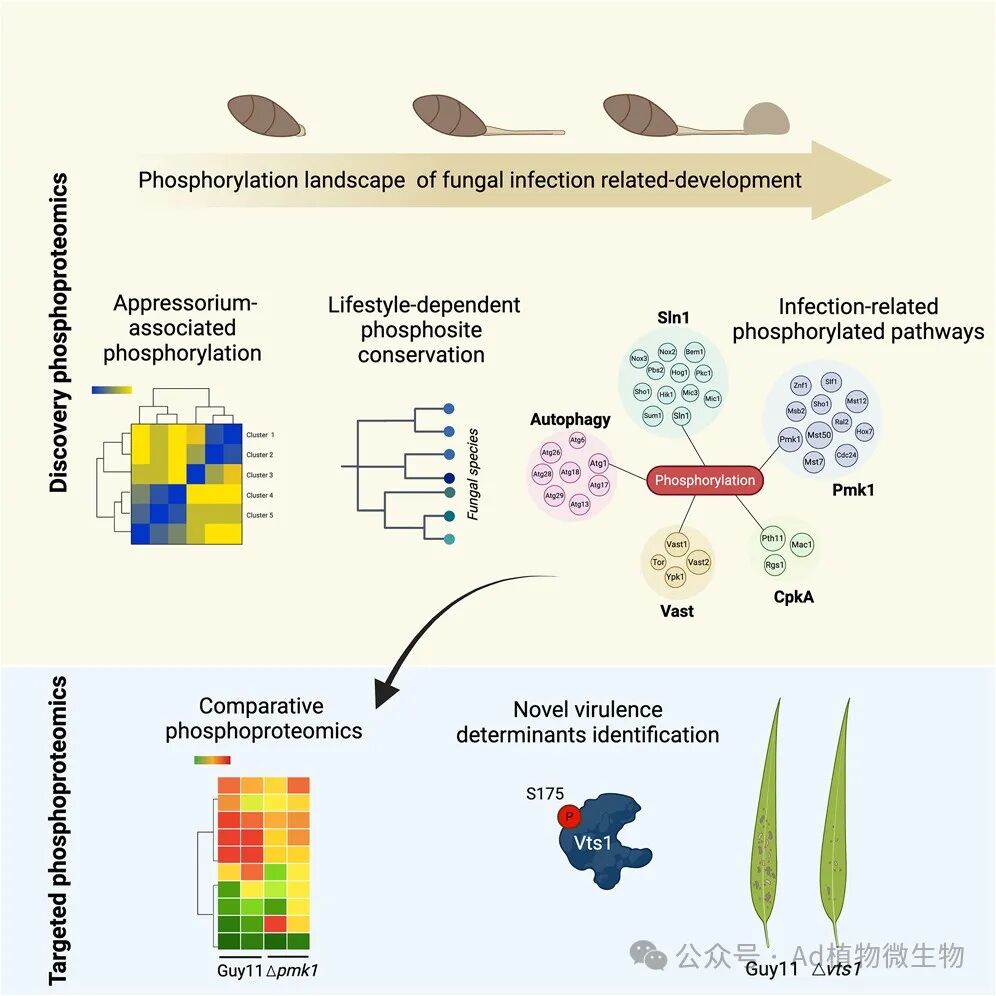
在这篇文章中,科研人员介绍了一种基于质谱法的定量磷酸化蛋白质组学分析,研究了威胁全球粮食安全的水稻稻瘟病菌的侵染相关发育过程。科研人员绘制了疏水表面萌发后 2,062 个真菌蛋白质上的 8,005 个磷酸化位点,揭示了附着胞发育过程中基于磷酸化的信号级联的主要重新布线。通过比较 41 种真菌的磷酸化保护情况,科研人员发现了与活体营养真菌和半活体营养真菌侵染特别相关的磷酸化特征。然后,科研人员利用平行反应监测(PRM)来识别受真菌 Pmk1 MAPK 调控的磷酸化蛋白,Pmk1 MAPK 可控制植物对 M. oryzae 的侵染。科研人员确定了 Pmk1 的 32 个底物,并证明稻瘟病需要调节因子 Vts1 依赖 Pmk1 的磷酸化。因此,定义侵染的磷酸化图谱可以确定控制植物病害的潜在治疗干预措施。